

Assorbimento ed efficienza d’uso dei nutrienti minerali
Le piante sono organismi autotrofi che ricavano il loro nutrimento da semplici fonti inorganiche: carbonio (C), idrogeno (H) e Ossigeno (O)
dall’aria e gli altri elementi sotto forma di cationi inorganici (K+) o ossidi (NO3-) dalla soluzione del terreno. Tutte le piante necessitano almeno di 16 elementi chimici e ciascuno di essi svolge funzioni biochimiche e fisiologiche specifiche nella cellula, tali da rendere questi elementi essenziali per la crescita e lo sviluppo della pianta.
Gli elementi nutritivi essenziali per la pianta vengono suddivisi in macro (se presenti nella pianta in quantitativi superiori a 1000 mg/Kg di Sostanza Secca “SS”) e micro nutrienti (se inferiori a 500mg/kg di SS).
Gli elementi contenuti nella pianta differiscono in quantità e qualità (non tutti sono essenziali, alcuni sono tossici) rispetto a quelli della litosfera (suolo), questo presuppone la presenza di un assorbimento selettivo. Le piante hanno quindi sviluppato meccanismi morfologici ed ecologici per garantirsi l’approvvigionamento di quantità sufficienti di elementi essenziali. Tali elementi vengono principalmente assorbiti dalla pianta a livello radicale (piccoli quantitativi anche a livello fogliare) e le loro mobilità nel suolo dipende da tre elementi: intercettazione radicale, flusso di massa e diffusione; il cui contributo varia in relazione al nutriente considerato. Per esempio, elementi quali N e Mg sono riforniti principalmente attraverso flusso di massa, mentre Potassio (K) e Fosforo (P) per diffusione.
L’efficienza di trasporto dei nutrienti, varia nelle diverse regioni costituenti l’apparato radicale. Per esempio, il nitrato in alcune specie, quali Mais e Citrus, viene maggiormente assorbito dalla zona apicale della radice. E’ stato dimostrato inoltre che la lunghezza delle radici laterali e la densità ed estensione dei peli radicali sono importanti fattori morfologici per l’acquisizione dei nutrienti per diffusione. Sebbene la relazione tra densità radicale e velocità di assorbimento non sia lineare, è indicativa che un’elevata densità diminuisce la velocità di assorbimento per effetto della sovrapposizione delle zone di esaurimento delle singole radici. La soluzione acquosa in cui sono contenuti gli elementi nutritivi, attraversa radialmente la radice mediante due vie principali: via apoplastica (flusso di massa e diffusione), simplastica (diffusione semplice, facilitata e trasporto attivo).
Gli elementi contenuti nella pianta differiscono in quantità e qualità (non tutti sono essenziali, alcuni sono tossici) rispetto a quelli della litosfera (suolo), questo presuppone la presenza di un assorbimento selettivo. Le piante hanno quindi sviluppato meccanismi morfologici ed ecologici per garantirsi l’approvvigionamento di quantità sufficienti di elementi essenziali. Tali elementi vengono principalmente assorbiti dalla pianta a livello radicale (piccoli quantitativi anche a livello fogliare) e le loro mobilità nel suolo dipende da tre elementi: intercettazione radicale, flusso di massa e diffusione; il cui contributo varia in relazione al nutriente considerato. Per esempio, elementi quali N e Mg sono riforniti principalmente attraverso flusso di massa, mentre Potassio (K) e Fosforo (P) per diffusione.
L’efficienza di trasporto dei nutrienti, varia nelle diverse regioni costituenti l’apparato radicale. Per esempio, il nitrato in alcune specie, quali Mais e Citrus, viene maggiormente assorbito dalla zona apicale della radice. E’ stato dimostrato inoltre che la lunghezza delle radici laterali e la densità ed estensione dei peli radicali sono importanti fattori morfologici per l’acquisizione dei nutrienti per diffusione. Sebbene la relazione tra densità radicale e velocità di assorbimento non sia lineare, è indicativa che un’elevata densità diminuisce la velocità di assorbimento per effetto della sovrapposizione delle zone di esaurimento delle singole radici. La soluzione acquosa in cui sono contenuti gli elementi nutritivi, attraversa radialmente la radice mediante due vie principali: via apoplastica (flusso di massa e diffusione), simplastica (diffusione semplice, facilitata e trasporto attivo).
Raggiunta l’endodermide, il trasporto diviene solo simplastico a causa della presenza della banda del Caspary che presenta suberina, sostanza altamente idrofoba. L’utilizzo di queste vie dipende dalle condizioni esterne, ed in particolare dalla concentrazione dei nutrienti. Per passare dall’apoplasto al simplasto, gli elementi nutritivi devono attraversare la membrana cellulare attraverso dei sistemi di trasporto altamente selettivi che discriminano i vari elementi in base alla carica ed al raggio ionico.
Alcuni elementi penetrano passivamente nella cellula attraverso processi di diffusione semplice o usufruendo di proteine di trasporto quali le acquaporine (specifiche per l’acqua) ed i canali che ne facilitano il movimento (diffusione facilitata). Specifici canali sono stati individuati per il trasporto del Ca++, K+, Na+, Cl- ed NO3-. Tuttavia i canali sono spesso chiusi ed il loro numero è piuttosto ridotto. Altri ioni invece vengono trasportati attivamente all’interno della cellula (trasporto attivo contro gradiente) e le proteine di trasporto dedite a questo compito sono chiamate carriers o pompe protoniche.
Alcuni elementi penetrano passivamente nella cellula attraverso processi di diffusione semplice o usufruendo di proteine di trasporto quali le acquaporine (specifiche per l’acqua) ed i canali che ne facilitano il movimento (diffusione facilitata). Specifici canali sono stati individuati per il trasporto del Ca++, K+, Na+, Cl- ed NO3-. Tuttavia i canali sono spesso chiusi ed il loro numero è piuttosto ridotto. Altri ioni invece vengono trasportati attivamente all’interno della cellula (trasporto attivo contro gradiente) e le proteine di trasporto dedite a questo compito sono chiamate carriers o pompe protoniche.
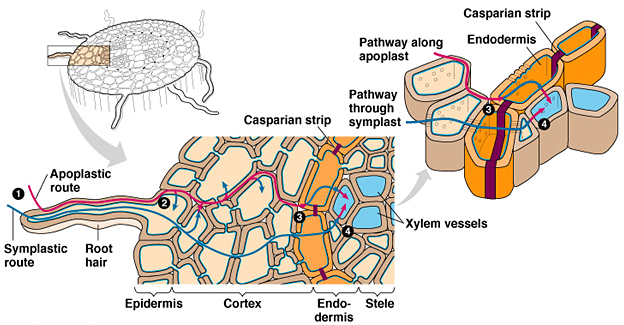
Immagine 1: Movimento della soluzione nutritiva all’interno dell’apparato radicale
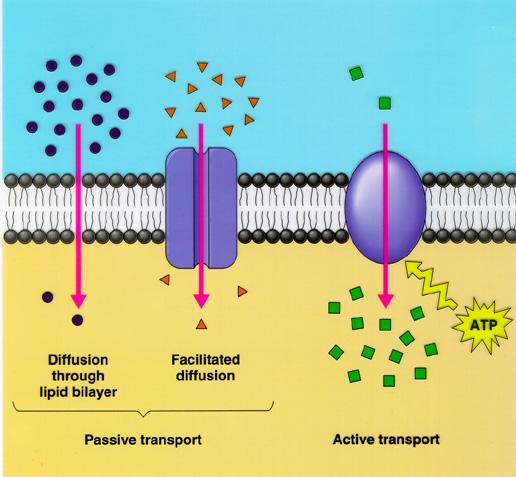
Immagine 2: Meccanismi di trasporto dei nutrienti attraverso la membrana cellulare
I carriers sono trasportatori proteici che presentano cinetiche saturabili rispetto alle variazioni di concentrazione esterne dello ione, e tali cinetiche sono simili a quelle che descrivono l’interazione enzima substrato. Questo significa che se abbiamo una pianta affamata nel momento in cui inizieremo a somministrargli dei nutrienti osserveremo ad un assorbimento esponenziale di questi fino ad un punto in cui tutte le molecole di trasporto sono saturate e l’assorbimento diventa lineare raggiungendo un plateaux. Quindi questo significa che se le piante sono ben nutrite una somministrazione di nutrienti superiore a quella saturante corrisponde ad una perdita di questi per mancato assorbimento. I carriers operano comunque un trasporto attivo secondario, poiché il trasporto attivo primario a livello di membrana plasmatica e tonoplastica è gestito dall’H+ ATPasi. La presenza di questi enzimi e la loro stretta relazione con il trasporto degli ioni e stata ormai accertata. Queste effettuando il trasporto elettrogenico di protoni creando un gradiente elettrochimico che rappresenta la forza protonica motrice che consente il trasporto secondario degli ioni che può avvenire tramite uniporto, sinporto o antiporto.
Superata la banda del Caspary, il trasporto di acqua e nutrienti avviene per flusso di massa, garantito dalla traspirazione. Uno dei parametri più importanti riguardanti la nutrizione è la NUE (Nutrient Use Efficiency). Le definizioni riguardanti la NUE sono numerose e spesso ambigue e possono essere espresse sia in termini agronomici (aspetti produttivi) che fisiologici (uso del nutriente).
Sono stati ricavati numerosi indici d’uso dei nutrienti che sono risultati spesso ambigui e contraddittori. Se si considera l’N si parla di efficienza d’assorbimento, N accumulato, concentrazione di N etc. Alcuni autori hanno proposto altri indici che permettono di distinguere i genotipi in efficienti e non efficienti e successivamente in superiori ed inferiori. Secondo tali autori il germoplasma efficiente ottiene una maggiore produzione rispetto a quello inefficiente, quando le condizioni nutrizionali sono limitanti, viceversa, in condizioni di buona disponibilità nutritiva, entrambe i germoplasmi raggiungono gli stessi livelli produttivi.
I germoplasmi invece, che ottengono diversa produttività anche in condizioni ottimali di disponibilità nutritiva, non possono essere comparati tra loro ed in tal caso si parla di genotipi superiori ed inferiori. Tuttavia non è sempre facile valutare i meccanismi anatomici, fisiologici e molecolari responsabili delle differenze d’efficienza d’uso dei nutrienti. Per esempio un genotipo con ampio apparato radicale è in grado di accumulare una maggiore quantità di fosforo (P) e migliorare le sue capacità produttive rispetto ad un altro con ridotto sviluppo. Nel caso dell’azoto (N) le differenze tra i genotipi, nell’efficienza d’uso di tale elemento, sono il prodotto di due componenti primarie: L’efficienza di assorbimento (capacità di estrarre l’elemento dal suolo), e l’efficienza d’uso del nutriente (capacità di convertire l’elemento assorbito nella produzione desiderata come ad esempio massa fogliare etc).
Per esempio, la capacità delle radici di assorbire nitrato dipende dalla sua concentrazione nel suolo, dal volume di suolo esplorato, dalla morfologia radicale (presenza di radici laterali e peli radicali) e dall’efficienza del sistema di trasporto. Quindi si può dire che i meccanismi che regolano l’espressione dell’efficienza d’uso dei nutrienti, in entrambe le sue componenti, possono essere di natura fisiologica, morfologica, anatomica, biochimica e molecolare. Per esempio in spinacio, una maggiore efficienza d’uso del nitrato, in zone povere di nutrienti, è correlata ad un incremento della superficie dei peli radicali.
L’efficienza di acquisizione di elementi quali il fosforo, può essere ottenuta anche attraverso la colonizzazione micorrizica delle radici che ne aumentano l’area superficiale. Ed infine, anche la capacità della pianta di utilizzare il nutriente accumulato al suo interno, gioca un ruolo essenziale nel migliorare l’efficienza nutritiva.
Sono stati ricavati numerosi indici d’uso dei nutrienti che sono risultati spesso ambigui e contraddittori. Se si considera l’N si parla di efficienza d’assorbimento, N accumulato, concentrazione di N etc. Alcuni autori hanno proposto altri indici che permettono di distinguere i genotipi in efficienti e non efficienti e successivamente in superiori ed inferiori. Secondo tali autori il germoplasma efficiente ottiene una maggiore produzione rispetto a quello inefficiente, quando le condizioni nutrizionali sono limitanti, viceversa, in condizioni di buona disponibilità nutritiva, entrambe i germoplasmi raggiungono gli stessi livelli produttivi.
I germoplasmi invece, che ottengono diversa produttività anche in condizioni ottimali di disponibilità nutritiva, non possono essere comparati tra loro ed in tal caso si parla di genotipi superiori ed inferiori. Tuttavia non è sempre facile valutare i meccanismi anatomici, fisiologici e molecolari responsabili delle differenze d’efficienza d’uso dei nutrienti. Per esempio un genotipo con ampio apparato radicale è in grado di accumulare una maggiore quantità di fosforo (P) e migliorare le sue capacità produttive rispetto ad un altro con ridotto sviluppo. Nel caso dell’azoto (N) le differenze tra i genotipi, nell’efficienza d’uso di tale elemento, sono il prodotto di due componenti primarie: L’efficienza di assorbimento (capacità di estrarre l’elemento dal suolo), e l’efficienza d’uso del nutriente (capacità di convertire l’elemento assorbito nella produzione desiderata come ad esempio massa fogliare etc).
Per esempio, la capacità delle radici di assorbire nitrato dipende dalla sua concentrazione nel suolo, dal volume di suolo esplorato, dalla morfologia radicale (presenza di radici laterali e peli radicali) e dall’efficienza del sistema di trasporto. Quindi si può dire che i meccanismi che regolano l’espressione dell’efficienza d’uso dei nutrienti, in entrambe le sue componenti, possono essere di natura fisiologica, morfologica, anatomica, biochimica e molecolare. Per esempio in spinacio, una maggiore efficienza d’uso del nitrato, in zone povere di nutrienti, è correlata ad un incremento della superficie dei peli radicali.
L’efficienza di acquisizione di elementi quali il fosforo, può essere ottenuta anche attraverso la colonizzazione micorrizica delle radici che ne aumentano l’area superficiale. Ed infine, anche la capacità della pianta di utilizzare il nutriente accumulato al suo interno, gioca un ruolo essenziale nel migliorare l’efficienza nutritiva.
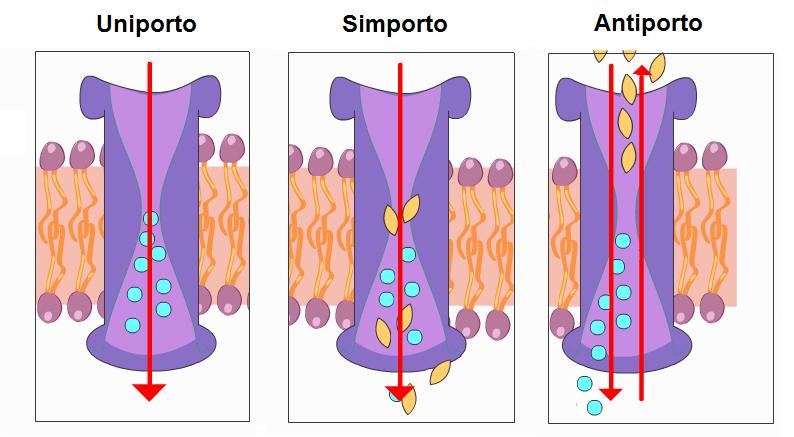
Immagine 3: Trasporto secondario degli ioni