

La scoperta delle citochinine
La scoperta delle auxine stimolò molti ricercatori nel cercare altri tipi di composti chimici capaci di regolare l’accrescimento vegetale,
in quanto sembrava improbabile che crescita e sviluppo delle piante fossero gestiti da una sola tipologia di ormoni. Questi studiosi focalizzarono i loro studi sulla ricerca di ormoni che fossero in grado di regolare la divisione cellulare.
I composti furono incontrati in tutte le parti della pianta, in particolare nei tessuti caratterizzati da un’attiva divisione cellulare come gli apici meristematici, nei semi in germinazione, nelle radici e nei frutti in accrescimento.
A questi fitoregolatori venne dato il nome di citochinine (da "citochinesi") ed è stato scoperto che queste molecole svolgono un ruolo fondamentale nella fasi successive alla replicazione del DNA ma antecedente alla divisione mitotica. Il primo studioso che propose l'esistenza di fattori che stimolano la divisione cellulare fu Wiesner nel 1892. Successivamente, nel 1913, Gottlieb Haberlandt scoprì che un composto trovato nel floema aveva la capacità di stimolare la divisione cellulare nelle cellule parenchimali.
A questi fitoregolatori venne dato il nome di citochinine (da "citochinesi") ed è stato scoperto che queste molecole svolgono un ruolo fondamentale nella fasi successive alla replicazione del DNA ma antecedente alla divisione mitotica. Il primo studioso che propose l'esistenza di fattori che stimolano la divisione cellulare fu Wiesner nel 1892. Successivamente, nel 1913, Gottlieb Haberlandt scoprì che un composto trovato nel floema aveva la capacità di stimolare la divisione cellulare nelle cellule parenchimali.
Ma fu solo nel 1941 che Johannes van Overbeek e successivamente F.C. Steward e collaboratori (1948) scoprirono che l'endosperma liquido di cocco addizionato ad una soluzione nutritiva era in grado di favorire la continua divisione di cellule adulte differenziate di uno svariato numero di specie portando alla formazione di callo. Questa scoperta mise in evidenza la presenza all’interno del latte di cocco di una sostanza o di un insieme di sostanze caratterizzate dalla capacità di stimolare l’entrata e la permanenza delle cellule mature nel ciclo di divisione cellulare.
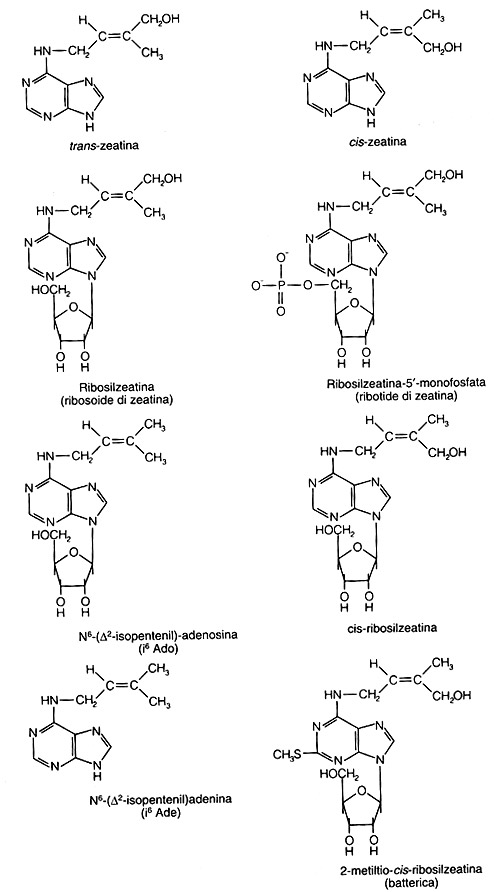
Successivamente si scoprì che all’interno del latte di cocco è contenuta la citochinina “zeatina”, ma tale scoperta venne fatta diversi anni dopo.
Nel 1954, Jablonski e Skoog dimostrarono che i tessuti vascolari contengono sostanze che stimolano la divisione cellulare e nel 1955, Folke Skoog e Miller Carlos testarono varie sostanze per la loro capacità di avviare e sostenere la proliferazione di cellule di tabacco e sorprendentemente scoprirono che autoclavando lo sperma di aringa riuscivano ad ottenere un prodotto, generato dal “break down” del DNA, che era in grado di promuovere e stimolare la divisione cellulare. La caratterizzazione chimica delle molecole portò alla scoperta della prima citochinina a cui venne dato il nome di “chinetina”.
La chinetina, che non esiste nelle piante, ha una struttura relativamente semplice ed i biochimici sono stati in grado di sintetizzare una varietà di altri composti analoghi che presentano la stessa attività biologica delle citochinine. Fu solo nel 1964 che Letham e colleghi furono in grado di isolare una citochinina naturale dai semi di mais (Zea mays), che prese il nome di zeatina (la stessa presente nell’endosperma di cocco), che è la citochinina naturale maggiormente attiva fra quelle note.
Effetti fisiologici prodotti dale citochinine
Sebbene siano state scoperte come un fattore di divisione cellulare, le citochinine sono in grado di stimolare e di inibire una serie di processi fisiologici, metabolici, biochimici e di sviluppo quando applicate alle piante superiori, ed è sempre più chiaro che le citochinine endogene svolgono un ruolo importante nella regolazione di questi eventi nella pianta. Fra queste attività ricordiamo la capacità di:
- indurre la divisione cellulare in cellule di callo in presenza di auxina (ormoni generalmente inseriti nei mastici cicatrizzanti per tagli giapponesi),
- promuovere la formazione di radici o germogli da colture in-vitro di callo quando utilizzate nei rapporti appropriati con le auxine,
- ritardare la senescenza fogliare e stimolano la mobilitazione delle sostanze nutritive e la sintesi della clorofilla,
- promuovere l'espansione dei cotiledoni fogliari nelle dicotiledoni,
- stimolare lo sviluppo delle gemme laterali e contrastare la dominanza apicale,
- stimolare la distensione cellulare e quindi l'espansione fogliare,
- Promuovere la conversione degli ezioplasti in cloroplasti attraverso la stimolazione della sintesi della clorofilla,
- Favorire la traspirazione fogliare aumentando il flusso idrico all’interno della pianta e quindi il trasporto dei nutrienti,
- Risvegliare le gemme dormienti ed eliminano la dormienza dei semi in alcune specie.
Le citochinine regolano il ciclo cellulare in radice e fusto, mentre il rapporto Auxina/Citochinina regola la morfogenesi in colture cellulari.
Studi sulle interazioni che coinvolgono auxina e citochinina stanno aiutando i fisiologi vegetali a capire come i fitormoni cooperino per produrre il modello di crescita complessivo di ogni pianta. Una cellula indifferenziata dispone di due opzioni: può estendersi, dividersi, estendersi nuovamente e ri-dividersi, o può semplicemente estendersi senza subire ulteriori divisioni. Le cellule che si dividono più volte rimangono sostanzialmente indifferenziate (cellule meristematiche), mentre le cellule che si distendono senza dividersi tendono a differenziasi, o specializzarsi andando a formare i tessuti. Studi effettuati su steli di tabacco micro coltivati misero in evidenza come l’aggiunta di IAA portavano ad una forte espansione delle cellule in coltura con la formazione di cellule giganti ma non alla divisione cellulare. L’aggiunta di sola chinetina non causava effetti degni di nota. Quando invece alla coltura cellulare veniva somministrata una miscela di IAA più chinetina si osservava la formazione di un gran numero di cellule relativamente piccole che permanevano nello stato indifferenziato. Da ciò è stato possibile intuire come auxine e citochinine collaborino nell’induzione della formazione di cellule meristematiche (apici vegetativi e radicali). Inoltre, in colture cellulari di callo, modificando il rapporto Auxina/Citochinina è possibile scegliere se favorire la formazione di radici o di steli. Infatti, una concentrazione più o meno uguale dei due ormoni, induce le cellule a rimanere indifferenziate (callo). Quando la concentrazione di auxina è più alta, dal tessuto indifferenziato si ha la formazione di radici. Di contro, con una maggiore concentrazione di citochinine si ha la formazione delle gemme. Equilibrando in maniera corretta i due ormoni si può avere contemporaneamente la formazione di entrambe gli organi e quindi la formazione di una piantina completa.
Le citochinine ritardano la senescenza
Un'altra caratteristica, apparentemente indipendente dalle citochinine è la capacità di prevenire la senescenza delle foglie. Nelle foglie staccate dalla pianta si osserva una lenta e continua degradazione della clorofilla, dell’RNA, dei lipidi e delle proteine, anche se tenute in ambiente umido ed arricchito di nutrienti. Questo processo di invecchiamento programmato che conduce alla morte è chiamato senescenza. I processi di invecchiamento fogliare sono più rapidi al buio che alla luce ed è stato osservato che il trattamento di foglie isolate con citochinine ne ritardano la senescenza. Se viene trattata solo una foglia, questa rimane verde anche dopo che le altre di simile età sono ingiallite e cadute. Anche il trattamento di una piccola parte del lembo fogliare porterà ad un ritardo della senescenza della parte trattata rispetto al resto della lamina.
A differenza delle foglie giovani in estensione, le foglie mature non sono in grado di produrre citochinine ed il ritardo del loro invecchiamento sarà dipendente dalla presenza delle citochinine derivate e trasportate dalle radici (zeatina riboside e diidrozeatina riboside) attraverso il flusso xilematico.
Le citochinine promuovono il movimento dei nutrienti
Le citochinine influenzano il movimento delle sostanze nutritive nelle foglie da altre parti della pianta. Questo fenomeno è noto con il nome di “mobilitazione dei nutrienti-citochinina indotta”. La dimostrazione di ciò è avvenuta somministrando dei nutrienti radioattivamente marcati e seguendo il loro spostamento nella pianta (tramite radiografia) dalle zone di accumulo alle zone trattate con le citochinine. I risultati ottenuti hanno messo in luce come i nutrienti siano preferenzialmente trasportati, ed accumulati nei tessuti trattati con le citochinine. Si è ipotizzato così che l'ormone provochi la mobilitazione dei nutrienti creando un nuovo rapporto “source-sink” (sorgente-pozzo).
Effetti fisiologici prodotti dale citochinine
Sebbene siano state scoperte come un fattore di divisione cellulare, le citochinine sono in grado di stimolare e di inibire una serie di processi fisiologici, metabolici, biochimici e di sviluppo quando applicate alle piante superiori, ed è sempre più chiaro che le citochinine endogene svolgono un ruolo importante nella regolazione di questi eventi nella pianta. Fra queste attività ricordiamo la capacità di:
- indurre la divisione cellulare in cellule di callo in presenza di auxina (ormoni generalmente inseriti nei mastici cicatrizzanti per tagli giapponesi),
- promuovere la formazione di radici o germogli da colture in-vitro di callo quando utilizzate nei rapporti appropriati con le auxine,
- ritardare la senescenza fogliare e stimolano la mobilitazione delle sostanze nutritive e la sintesi della clorofilla,
- promuovere l'espansione dei cotiledoni fogliari nelle dicotiledoni,
- stimolare lo sviluppo delle gemme laterali e contrastare la dominanza apicale,
- stimolare la distensione cellulare e quindi l'espansione fogliare,
- Promuovere la conversione degli ezioplasti in cloroplasti attraverso la stimolazione della sintesi della clorofilla,
- Favorire la traspirazione fogliare aumentando il flusso idrico all’interno della pianta e quindi il trasporto dei nutrienti,
- Risvegliare le gemme dormienti ed eliminano la dormienza dei semi in alcune specie.
Le citochinine regolano il ciclo cellulare in radice e fusto, mentre il rapporto Auxina/Citochinina regola la morfogenesi in colture cellulari.
Studi sulle interazioni che coinvolgono auxina e citochinina stanno aiutando i fisiologi vegetali a capire come i fitormoni cooperino per produrre il modello di crescita complessivo di ogni pianta. Una cellula indifferenziata dispone di due opzioni: può estendersi, dividersi, estendersi nuovamente e ri-dividersi, o può semplicemente estendersi senza subire ulteriori divisioni. Le cellule che si dividono più volte rimangono sostanzialmente indifferenziate (cellule meristematiche), mentre le cellule che si distendono senza dividersi tendono a differenziasi, o specializzarsi andando a formare i tessuti. Studi effettuati su steli di tabacco micro coltivati misero in evidenza come l’aggiunta di IAA portavano ad una forte espansione delle cellule in coltura con la formazione di cellule giganti ma non alla divisione cellulare. L’aggiunta di sola chinetina non causava effetti degni di nota. Quando invece alla coltura cellulare veniva somministrata una miscela di IAA più chinetina si osservava la formazione di un gran numero di cellule relativamente piccole che permanevano nello stato indifferenziato. Da ciò è stato possibile intuire come auxine e citochinine collaborino nell’induzione della formazione di cellule meristematiche (apici vegetativi e radicali). Inoltre, in colture cellulari di callo, modificando il rapporto Auxina/Citochinina è possibile scegliere se favorire la formazione di radici o di steli. Infatti, una concentrazione più o meno uguale dei due ormoni, induce le cellule a rimanere indifferenziate (callo). Quando la concentrazione di auxina è più alta, dal tessuto indifferenziato si ha la formazione di radici. Di contro, con una maggiore concentrazione di citochinine si ha la formazione delle gemme. Equilibrando in maniera corretta i due ormoni si può avere contemporaneamente la formazione di entrambe gli organi e quindi la formazione di una piantina completa.
Le citochinine ritardano la senescenza
Un'altra caratteristica, apparentemente indipendente dalle citochinine è la capacità di prevenire la senescenza delle foglie. Nelle foglie staccate dalla pianta si osserva una lenta e continua degradazione della clorofilla, dell’RNA, dei lipidi e delle proteine, anche se tenute in ambiente umido ed arricchito di nutrienti. Questo processo di invecchiamento programmato che conduce alla morte è chiamato senescenza. I processi di invecchiamento fogliare sono più rapidi al buio che alla luce ed è stato osservato che il trattamento di foglie isolate con citochinine ne ritardano la senescenza. Se viene trattata solo una foglia, questa rimane verde anche dopo che le altre di simile età sono ingiallite e cadute. Anche il trattamento di una piccola parte del lembo fogliare porterà ad un ritardo della senescenza della parte trattata rispetto al resto della lamina.
A differenza delle foglie giovani in estensione, le foglie mature non sono in grado di produrre citochinine ed il ritardo del loro invecchiamento sarà dipendente dalla presenza delle citochinine derivate e trasportate dalle radici (zeatina riboside e diidrozeatina riboside) attraverso il flusso xilematico.
Le citochinine promuovono il movimento dei nutrienti
Le citochinine influenzano il movimento delle sostanze nutritive nelle foglie da altre parti della pianta. Questo fenomeno è noto con il nome di “mobilitazione dei nutrienti-citochinina indotta”. La dimostrazione di ciò è avvenuta somministrando dei nutrienti radioattivamente marcati e seguendo il loro spostamento nella pianta (tramite radiografia) dalle zone di accumulo alle zone trattate con le citochinine. I risultati ottenuti hanno messo in luce come i nutrienti siano preferenzialmente trasportati, ed accumulati nei tessuti trattati con le citochinine. Si è ipotizzato così che l'ormone provochi la mobilitazione dei nutrienti creando un nuovo rapporto “source-sink” (sorgente-pozzo).